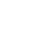

鄱阳湖重点实验室最新研究为白鹤种群的遗传拯救提供新见解
时间:2024-12-20 来源:
论文信息
南昌大学鄱阳湖环境与资源利用教育部重点实验室为第一单位的研究成果近期以“Understanding the Past to Preserve the Future: Genomic Insights Into the Conservation Management of a Critically Endangered Waterbird”为题发表在国际分子生态学经典期刊Molecular Ecology上,北京林业大学生态与自然保护学院王文娟副教授和中山大学生态学院刘阳教授为通讯作者,南昌大学鄱阳湖环境与资源利用教育部重点实验室已毕业硕士生,中山大学在读博士生陈青为论文第一作者,中山大学刘阳课题组博士生林洪洲、郑晨晴博士,俄罗斯科学院Elena A. Mudrik,俄罗斯奥卡鹤类繁育中心Tatiana A. Kashentseva,河北大学生命科学学院程亚林研究员,江西省林业科学研究院汪志如研究员,南昌五星白鹤保护小区周海燕,沈阳理工大学环境与化工学院周海翔教授也参与了本项工作。该研究得到了国家自然科学基金(32160132,32471732)和中央高校基础科研基金(BLX202252)的资助。

研究背景
白鹤(Leucogeranus leucogeranus)被世界自然保护联盟(IUCN)列为“极危”(CR)物种,为我国一级重点保护动物。在地理上分为东部(EA)和西部/中部(WCA)两个种群(图1)。东部种群主要在俄罗斯东部繁殖,迁徙至鄱阳湖越冬。东部种群在1981年之前并未有其记录,直到1981在鄱阳湖发现100多只个体,随后数量逐年增长,现今已接近7000只。西部/中部种群主要在俄罗斯西部繁殖,迁徙至伊朗和印度越冬,其种群数量在近几十年迅速下降,现已无野外个体的记录。为了恢复西部/中部种群,由世界鹤类基金会(ICF)和前苏联的鸟类学家共同建立了奥卡鹤类繁殖中心(OCBC),该中心从1976年开始开展人工繁殖和野外种群再引入工作。在该繁育中心圈养的白鹤主要由东部和西部/中部种群繁殖地获取的鸟卵孵化而来,当时收集了5枚西部/中部种群的鸟卵,目前有4只个体依然存活,1只个体已死亡。迄今为止,奥卡鹤类繁殖中心释放了近200只人工繁育个体至西部/中部种群的迁飞路线上,但是基本都死亡或者杳无音讯。为了防止该种群的灭绝,重建西部/中部种群的工作迫在眉睫。
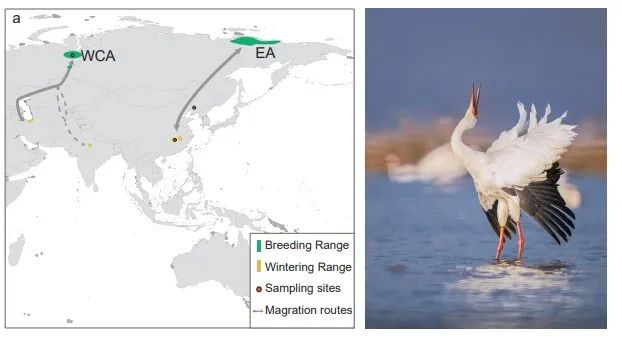
图1. 白鹤地理分布及采样点
圈养个体再引入是恢复西部/中部种群主要的解决方案。在濒危物种再引入过程中,常常会将不同种群进行交配,这有助于提高物种的遗传多样性、减轻近亲繁殖的负面影响、增强物种的进化潜力,从而促进种群的恢复(也被称为遗传拯救)。然而,当遗传差异较大的种群进行交配时,也存在远交衰退的风险(由于共适应基因复合体的破坏或不适应等位基因的增加导致适合度下降的现象)。因此,需要科学评估不同繁殖种群的近亲繁殖和远交衰退风险,从而为濒危物种的遗传管理和再引入工作的开展提供科学依据。
目前,针对濒危物种的遗传管理和再引入工作,大都主要关注近交衰退的风险,对于远交衰退的关注度较低,这主要是由于远交衰退不是导致物种灭绝的常见原因,并且其评估较为困难。目前已有的评估远交衰退的方法主要基于推荐迁移决策树。该评估框架考虑了以下四个标准:(1)明确的分类地位(种群结构);(2)没有固定的染色体差异;(3)具有较近期的隔离(<500年);(4)栖息环境相似。在满足这些标准时实施遗传拯救,远交衰退风险较低,对物种的适合度具有积极影响。对远交衰退的担忧可能会限制对濒危物种的一些管理行动,因此需要同时对近亲繁殖和远交衰退的风险开展科学评估。
主要发现
为了评估使用白鹤东部种群对西部/中部种群进行遗传拯救的风险,由中国和俄罗斯鸟类学家组成的研究团队联合开展了白鹤保护基因组学研究。通过对17个个体(12个东部个体和5个西部/中部个体)的重测序数据,评估了东部和西部/中部群体之间的遗传分化、种群历史动态、基因流以及遗传负荷。研究结果表明白鹤东部和西部/中部群体形成了不同的遗传分支,但它们之间的遗传分化程度较低(图2)。白鹤在更新世经历了有效种群大小剧烈波动,这可能是由于大规模的气候波动和环境变化所引起。大约在3.3万年前两个种群开始分化,并且有较为近期的基因流发生(图3)。
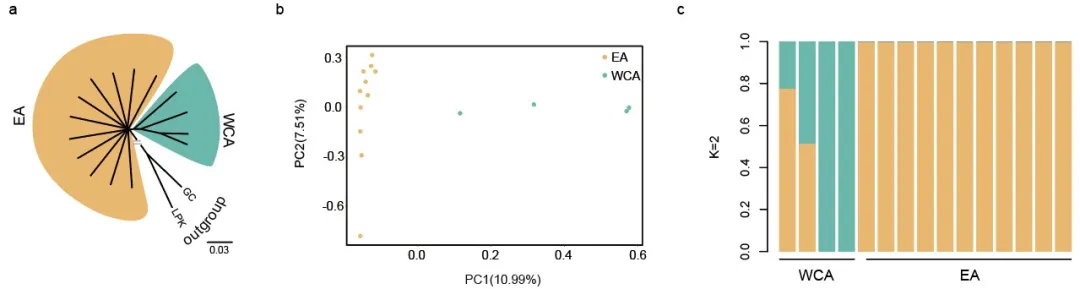
图2.白鹤的遗传结构。(a)系统发育树 (b)主成分分析(c)Admixture分析(k=2)
图3.种群历史动态。(a)PSMC重建古老历史 (b)GONE评估近期历史 (d)fastsimcoal模拟种群历史动态
研究结果同时表明,长期的种群下降导致两个种群低水平的遗传多样性和高水平的近亲繁殖(图4)。两个种群之间的杂合度没有显著差异(图4a)。相比于其他鸟类杂合度,白鹤的杂合度偏低(图4b),表明其对未来环境变化的适应潜力较弱。两个种群的近亲繁殖大都发生于较远时期(图4c),西部/中部种群的近亲繁殖程度高于东部种群(图4d和4e),且近期的种群瓶颈效应更为严重(图4f)。
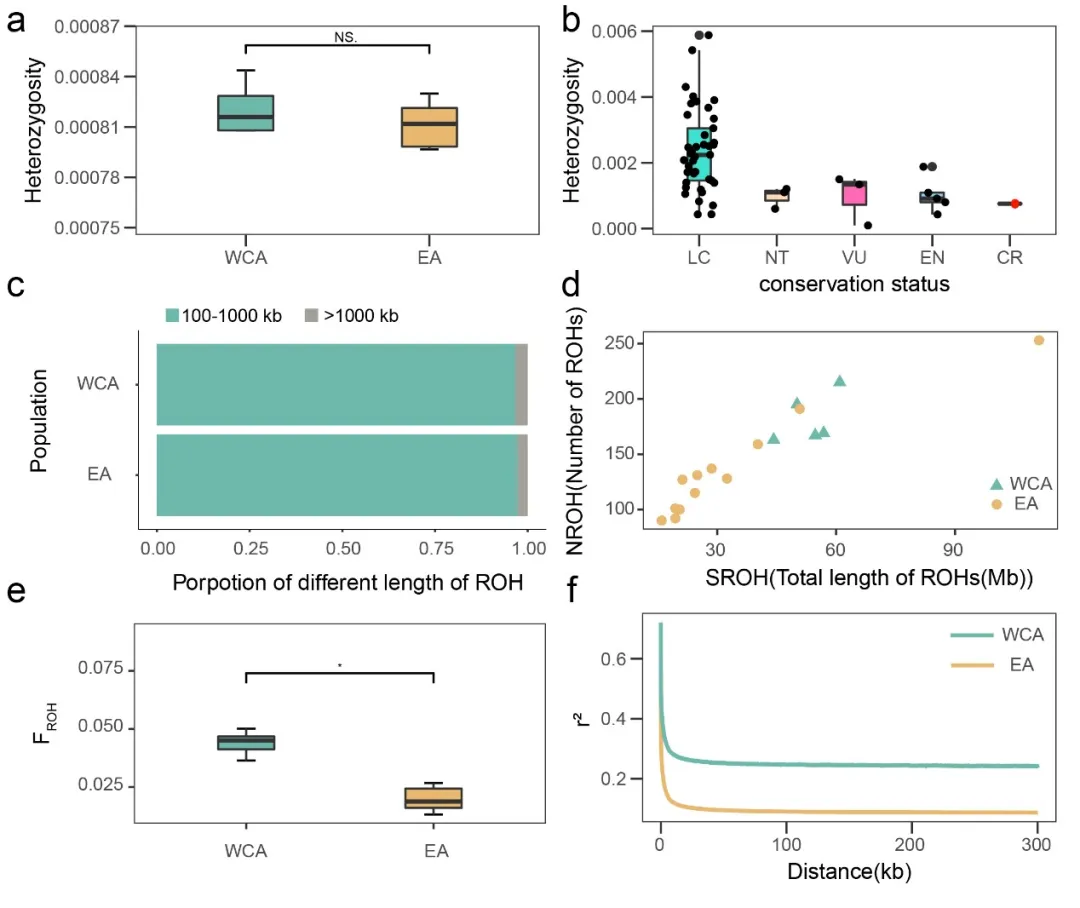
图4.遗传多样性和近亲繁殖水平。(a)杂合度 (b)已发表鸟类物种的杂合度 (c)ROH长度比例 (d)ROH片段的总长度和数量 (e)近交系数(FROH) (f)连锁不平衡衰减
西部/中部种群遭受着近交衰退的负面影响(图5)。尽管部分功能丧失突变得到了清除,但仍有过多的有害错义突变,且ROH区域的有害突变并未被显著移除(图5d,e,f)。模拟结果进一步表明西部/中部种群在种群数量迅速下降的过程中遭受了严重的近交衰退,适应度明显下降。
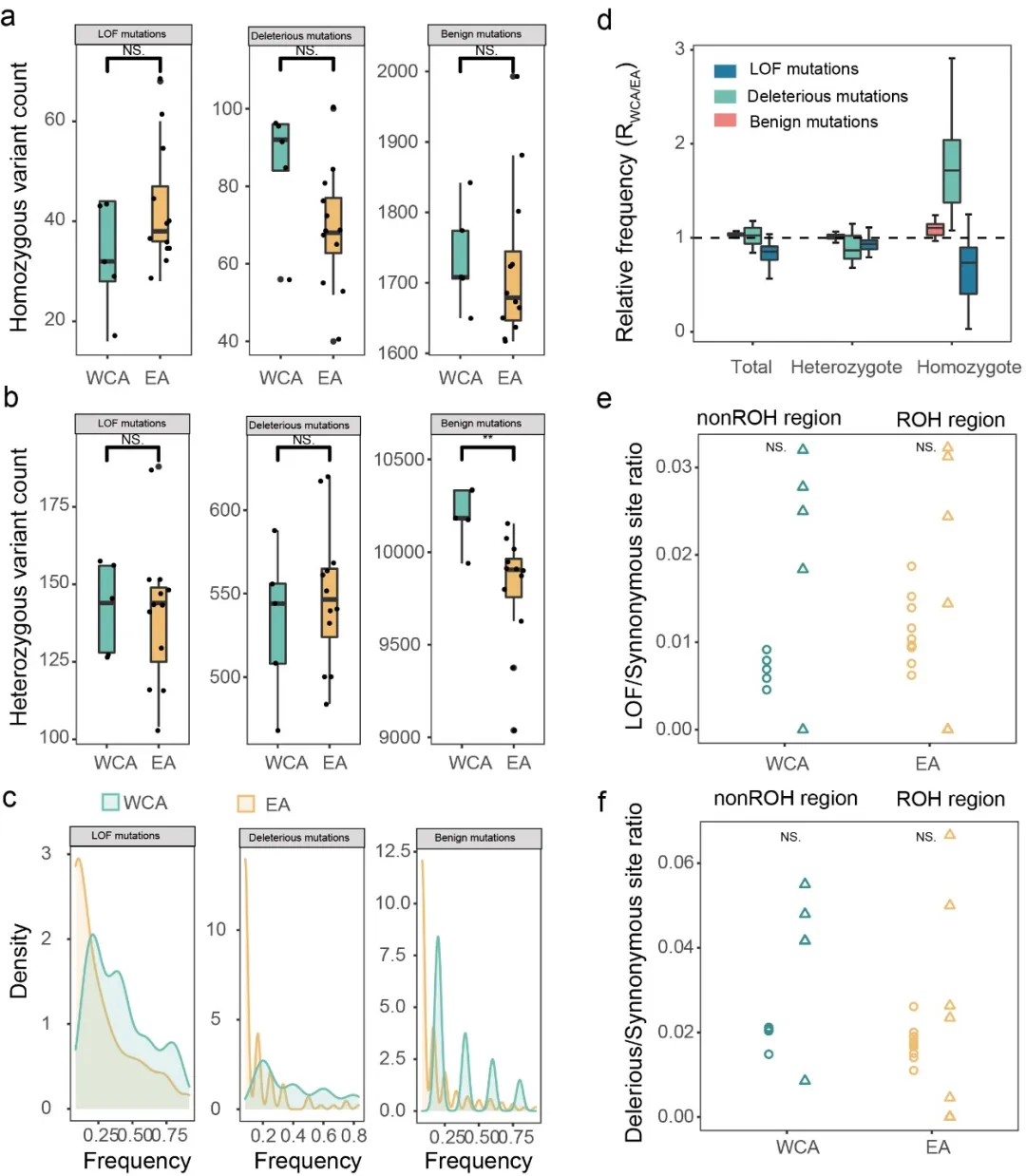
图5.遗传负荷。(a,b)功能丧失突变(LOF)、有害错义突变和良性错义突变的纯合子和杂合子数量(c)三种纯合变异的频率和密度(d)三种突变的纯合和杂合位点的相对频率 (e,f)ROH区域和非ROH区域中功能丧失突变和有害错义突变与同义突变的比值
东部种群和西部/中部种群之间较小的遗传差异及一定的种群间基因流表明,利用东部个体作为源种群与西部/中部种群交配的远交衰退风险较低。尽管如此,在将东部个体和西部/中部个体杂交时,还需要考虑局部适应和有害突变的潜在风险。本研究表明两个种群的差异基因主要与疾病发生相关,而与环境适应性特征的关联并不显著。同时,在进行杂交时,也需要考虑可能引入的有害突变。总体而言,本研究为濒危物种的遗传管理前的科学评估提供了一个示例,并强调了利用基因组方法评估远交衰退可能性的重要性。
论文链接:https://doi.org/10.1111/mec.17606